CA-MRSA: Background, Introduction and Singapore
The Singapore General Hospital Diagnostic Bacteriology Section blogger has just posted a succinct and comprehensive overview of community-associated MRSA (CA-MRSA) clones in Singapore. It seems like ancient history now, but CA-MRSA really rose into the global consciousness only in the early 21st century, when scientists from Europe (particularly the researchers from INSERM E0230, Lyon France – the Centre National de Référence Staphylocoques), U.S.A. and Australia published a joint paper in Emerging Infectious Diseases showing the different molecular characteristics that defined CA-MRSA at that time – namely, the presence of the Panton-Valentine Leukocidin and the presence of staphylococcal cassette chromosome mec type IV (SCCmec IV).
It is probably worthwhile to provide a more detailed description of these genetic elements here:
The history of the elegantly-sounding Panton-Valentine leukocidin (PVL) is well described in this article by authors from INSERM E0230. Although some Staphylococcus aureus strains were known to be “leukotoxic” (i.e. destroyed white blood cells) since 1894, it was only in the mid-1930s that Philip Noel Panton (he certainly had a colourful career as can be seen from his obituary here) and Francis Valentine from the Hale clinical laboratory at London Hospital isolated the toxin, which was subsequently named after them in 1936. And it might well have been consigned to the bin of esoteric bacterial toxins had it not been for its two signal associations:
- S. aureus strains with the PVL toxin were isolated almost exclusively from two types of infections only – furuncles/cutaneous abscesses and severe necrotising pneumonia.
- The majority of “true” CA-MRSA carry the genes for the PVL toxin – this association is so far unexplained. The PVL genes are carried on two distinct phages (viruses that target bacteria), which is presumably how they are passed between different S. aureus strains, but what this has to do with acquisition of SCCmec IV (or others) is unclear.
SCCmec is the mobile genetic element that carries the mec gene, which confers “methicillin resistance” on the bacteria by coding for a penicillin-binding protein (PBP2a) with reduced affinity for virtually all beta-lactam antibiotics (i.e. penicillins, cephalosporins and carbapenems). We have finally overcome this barrier after 50 years, with “5th generation cephalosporins” created (such as ceftobiprole, ceftaroline and ceftolozone) that are able to bind to PBP2a. However, their efficacy in serious MRSA infections has not been rigorously tested yet. There are multiple different types of SCCmec in circulation (last count is 11, according to the website of the International Working Group on Staphylococcal Chromosome Cassette Elements). SCCmec is not native to S. aureus, but probably originated from other staphylococci, including possibly those of animal origin. Up to the mid-1990s, there was only SCCmec I-III, but SCCmec IV soon appeared in CA- and subsequently HA-MRSA from the mid-1990s, followed by SCCmec V, after which perhaps a more intense search and better sequencing technology resulted quickly in the discovery of the other SCCmec types.
But back to CA-MRSA.
A historical timeline of its emergence is provided here by the MRSA Research Center at the University of Chicago Medicine. CA-MRSA was first noticed in the late 1980s in the aboriginal population in Western Australia and also in parts of the U.S.A. The tools for identifying and typing CA-MRSA (SCCmec, MLST, PVL) were not available then, and I recall listening to a lecture by Prof Robert Daum years ago where he described the difficulties he had in publishing the initial paper on CA-MRSA in the U.S.A. A reviewer had rejected his initial submission with a short sentence “there is no such thing as CA-MRSA!”. It was not until 1997-1999, where four young children in Minnesota and North Dakota died of severe infection caused by CA-MRSA, that the concept of CA-MRSA as a separate entity (rather than just “feral” healthcare-associated MRSA) started to gain traction.
In the first decade of the 21st century, large-scale CA-MRSA epidemics occurred in several regions and countries of the world, including U.S.A., Western Australia, Taiwan, Hong Kong, parts of Europe, and Algeria. CA-MRSA infections are a notifiable disease in Hong Kong, whereas in the other countries/regions described above, CA-MRSA rates were so high that the bacteria invaded the hospitals, replacing the endemic HA-MRSA in some cases and causing healthcare-associated infections.
Several groups had looked out for CA-MRSA in Singapore too. Prof Paul Tambyah’s group at the National University Hospital relied entirely on epidemiological criteria, and found that only one of 383 MRSA infections during a one-year period had occurred in a patient with no significant healthcare contact. Our own work, using molecular techniques to prove that the isolate in particular was a CA-MRSA strain, has been described by the SGH Diagnostic Bacteriology blog.
This work at SGH took place almost purely because of a string of happy coincidences. I had written to Prof Jerome Etienne at the end of 2003 – then director of INSERM E0230 – to discuss the case of PVL necrotising pneumonia in Singapore and to see if I could visit his laboratory. Much to my surprise, I was warmly welcomed (he didn’t know me from Adam) and spent a pleasant two days in March 2004 at INSERM E0230 in Lyon, meeting with other prominent researchers (including those who had described the initial association between PVL and necrotising staphylococcal pneumonia), joining in an ICU ward round (like much of Europe, France does not have many ID physicians and much of that work is done by clinical microbiologists) and learning their protocols. There was certainly no overt secrecy and they freely shared their protocols with me (two printed sheets of PCR primer sequences and the PCR thermal cycling sequence), as well as findings on CA-MRSA.

View of Lyon, France. Unfortunately, my better photos from 2004 are lost.
Back in Singapore, it turned out that A/Prof Koh Tse Hsien from SGH Diagnostic Bacteriology Section had kept a number of MRSA isolates that had different susceptibility profiles compared to the usual ST239-MRSA-III that was dominant in our hospitals then. Five of these isolates belonged to patients who had not obviously been exposed to our healthcare system previously, based on their medical records (the rest, as it turned out, were UK-EMRSA-15 isolates). Using the protocols freshly obtained from INSERM E0230 in Lyon, we were able to show that these were all PVL-positive (below), and two isolates had SCCmec IV, whereas three isolates had untypeable SCCmec (SCCmec V was described shortly after this work and we confirmed that these were indeed SCCmec V). The researchers from Lyon helped with the MLST and re-confirmed the toxin gene profiles. Elated by our “success” in finding local CA-MRSA, we continued with our local surveillance for a number of years before concluding our efforts in early 2008 due to a combination of bureaucratic and financial factors.
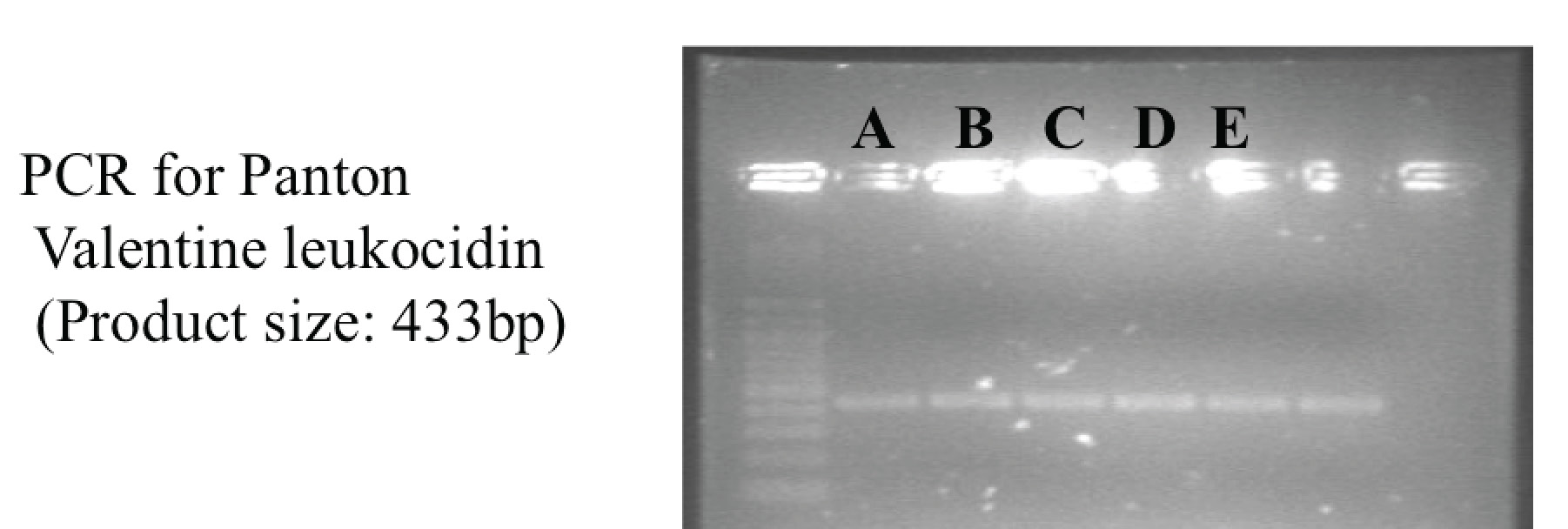
Gel electrophoresis showing positive PCR products for Panton-Valentine Leukocidin for the 5 initial CA-MRSA isolates in Singapore.
Unfortunately, after 2008, there has not been a concerted effort to study the problem of CA-MRSA in Singapore. Anecdotally, the rates have been increasing in various local hospitals, but not to the extent as in other countries where these have started to replace HA-MRSA. One of the incidental findings of our survey of MRSA carriage isolates for presence of mupirocin and ACME (published in the Journal of Global Antimicrobial resistance – behind a paywall unfortunately) was that approximately 4.5% (30 isolates of 666) were CA-MRSA, of which a slight majority belonged to ST772 (the most common CA-MRSA clone in India).
